Articles by Author | Articles by Topic
The Bions: An Investigation into the Origin of Life
Wilhelm Reich, M.D.
Reprinted from the Journal of Orgonomy, Vol. 10 No. 1
The American College of Orgonomy
Editorial Note:
The Journal is proud to present the first English translation of Die Bione, a landmark monograph first published in Oslo in 1938 and still, today, decades ahead of its time.
The momentousness of Reich's discoveries set forth here cannot be over-estimated. While the scientific world still clung to the dictum that "every living thing comes from living matter," Reich began to probe the origins of life from a functional point of view. He has already enunciated his orgasm formula of tension-charge-discharge-relaxation as specific to all living phenomena and encompassing every aspect of the autonomic life functions from ameba to man. In a series of human experiments, he had shown empirically the existence of an organismic energy (which he still thought of as "bioelectric" energy) and had proved that the subjective feelings of pleasure and anxiety were nothing other than one's perceptions of this objective, measurable energy, as it flowed from the core to periphery and periphery to core, respectively. This to-and-fro movement of energy represented the basic life function of pulsation (contraction-expansion), mediated through respiration in man and experienced as pleasure or anxiety whenever a certain energy threshold was reached.
Going from macrocosm to microcosm, Reich now ventured into the riddle of biogenesis. His intent was to set up processes in various substances that would replicate the four-beat fundamental formula. He found that both organic and inorganic matter, when made to swell, disintegrated into fluid-filled vesicles charged with energy and visible only at magnifications upwards of 2000x. Preparations were first sterilized or autoclaved at critical temperatures to rule out the presence of prior life forms. The resulting vesicles gave off a bluish glimmer and showed such properties as pulsation, locomotion, internal vibration, Gram-positive reaction, division, fusion, and the ability to paralyze and destroy bacteria and small protozoa. Reich described his discovery as "a minute quantity of matter, containing a quantity of energy derived from the matter…not complete living beings but only carriers of biological energy…forms of transition from non-living to living." He called the vesicles "bions" and spent many hours observing them and taking time-lapse films. To accomplish this required a virtuoso grasp of microscope technique and mastery of a thousand technical problems. A monumental film (which this writer has seen and which now reposes in the Reich archives) shows the bionous disintegration of a grass infusion and its subsequent reorganization into protozoa. This led inevitably to his discovery of an identical process in the formation of cancer cells and to the historic Experiment XX, the natural organization of protozoa from a sterile solution of bion water which had been autoclaved, frozen, and then thawed.
Reich was quick to caution that he had not created life from the non-living, but had demonstrated "the natural process by which protozoa and cancer cells develop spontaneously from vesicularly disintegrated matter." It took but one giant step for Reich to realize that the energy emanating from the bions was the same "bioelectricity" found in all living organisms from man to ameba, that it permeated everything and was cosmic in nature. In one stroke, orgone energy, as he now called the cosmic life energy, emerged from the shadowy domain of man's intuition and entered the realm of scientific investigation.
BGK
Introduction
It is not without concern that I make public my current experimental results on the origin of the vegetative life. This has nothing to do whatsoever to do with the correctness and accuracy of the data, though an insignificant error or inept wording might well be found here and there. There are no results in this comprehensive, if not final, report that have not been confirmed hundreds of times. Observations that could not be verified were omitted. I have tried to describe the experimental method as clearly as possible in order to render replication feasible. The basic facts, such as the vesicular disintegration of matter due to swelling, or the cultivation of bions, are inescapable when the directions are followed with some measure of accuracy. I am aware that one can explain these very facts in a way that differs from my own interpretation. For this reason, I have carefully separated the data section in the first part from the interpretation in Part Two.
I am concerned about the possibility of being branded presumptuous in my ensuing hypothesis. I did not step outside the framework of reference laid down by eighteen years of clinical work on the vegetatively sick organism and ten years of intensive study of the pertinent literature in biology and physiology. The sections dealing with colloids and the dialectic materialistic method of investigation were in my desk drawer for many years, completed but unpublished. They were attempts to correlate my clinical experiences as a psychotherapist with my general biological studies. When I had to review Kraus's book on human pathology (Syzygiologie) for a scientific journal in 1926, I immediately recognized, on the basis of my orgasm theory, its tie-in with psychoanalytic knowledge.
I had no idea that ten years later I would be able to verify hypotheses in natural science and methodology of dialectic materialism to such an extent. I knew, of course, that the orgasm theory pointed toward the problem of organic life. We are not dealing with an accidental discovery, but a long-term development of work on the problem of vegetative life functions. The foundation for a theory of biogenesis, slated to unfold subsequently, was uncovered step by step. I must admit that the facts I came across seemed unbelievable to me at first. However, one fact followed another. Every single one confirmed the idea that had come to me previously in my work with patients concerning the life functions and their disturbances. At the time I published the experimental data on the electrical function of sexuality and anxiety in 1937. [Footnote 1]
I already had in my possession the cultures resulting from my bion research. And now that I have decided to release this material for publication, I have already obtained further data in a related field, which confirm and amplify the previous results.
The technique employed in the experiments does not differ from the usual bacteriological sterilization procedures. However, the experimental setup, my way of thinking things through, and my interpretations diverge considerably from the usual. The experimental format was always geared to the basic sexual formula that I discovered in my research on sexuality. Thought processes follow laws that are consistent with dialectic materialism. The doctrine set forth by Hegel, which Marx had extended to materialism and Engels had first applied to the natural sciences, now found new application in psychology and in sexual functioning. Many of its theories were thereby strengthened. New insights were gained: for example, the formulation of the "dialectic materialistic doctrine of evolution." From Freud I took over the idea of equating life drives and sexual drives. After I had succeeded in refuting his theory of the death instinct and in developing the orgasm theory, the door was open to experimental biology. Consequently, the experimental evidence for the identity of sexual energy and life energy processes is at the same time a confirmation of Freud's hypothesis.
I am especially grateful to Professor Roger du Teil for the invaluable friendship he extended in this endeavor. Whatever the outcome of his efforts to steer the world of biology and bacteriology toward this work, his active collaboration in the research had become an organic part of the whole. It is clearly apparent in this paper.
I am also aware that the experimental solution to the question of spontaneous generation only partially fills the hiatus existing throughout the scientific world. Hence, there is going to be strong opposition. However, evidence, counter-evidence, and new evidence are part of the essence of scientific work. Furthermore, every objection begets progress if the fundamental problem is approached correctly.
I set forth the historical development of the problem in my paper "Dialectical Materialism in the Investigation of Life Functions" (Zeitschr. F. pol. Psych. U. Sexok., H. 3, Bd. IV, 1937). It also reviews the relevancy to sociological questions. I postponed a detailed discussion of many studies and follow-up questions pending until later publications.
Professor Harald Schjelderup merits great credit for having made possible and actively promoting the preliminary electrophysiological experiments at his university's Institute of Psychology. Without his overall efforts, more difficulties would have piled up.
We grappled with tremendous financial problems in setting up the entire laboratory. It would not have been possible to perform the experiments at another otherwise occupied institution. I could never have done it by myself. The Rockefeller Foundation in Paris had refused to give any support. Therefore, my special and heartfelt thanks to all those friends who pushed through the difficult birth of this project: in particular, my friend Sigurd Hoel, whose advice often prevented me from losing faith in this project; our friend Dr. Odd Havrevold, who equipped the research laboratory, lent a helping hand everywhere, and obtained funds; my assistants in the bacteriological, micro film. And physiochemical work, whose initiative helped me carry on through many vicissitudes. Much could not have been accomplished without active financial support from the Institute through my colleagues in the characteranalytic field; they helped to engineer the entire setup, and they remained loyal: Dr. Lotte Liebeck; Dr. Nic Hoel; Dr. Ola Raknes; Dr. Tage Philipson; Dr. Leunbach; Ellen Siersted.
The efforts of people who themselves did not possess any funds would not have been sufficient. Large contributions from Mr. Lars Christensen (Oslo), and Mr. Rolf Stenersen (Oslo), as well as Constance Tracey (London) were crucial in supporting the endeavor.
The biological laboratory alone required about 60,000 Norwegian krone for equipment. At the moment, the operation costs about 2,000 krone per month.
The whole work was greatly aided by the efficiency of my office workers, especially my secretary Gertrud Brandt, who tirelessly tended the orderly handling of my extensive activities. The bookkeeping was done by our publication manager Harry Proll, whose painstaking care is readily apparent.
The Institute was founded by Norwegians. The superlative Norwegian hospitality provides the fertile territory and foundation for my work, for whose leadership I assume full responsibility: Norway, a country that basically still knows how to hold at bay the mental anguish of the world.
The Major Items of Laboratory Equipment
(See: Plates I-VI.)
The complex experiments for assessing the electro- and micro-biological properties of the diverse bions and substances required special adaptation, and, in some cases, specially purchased equipment.
The Microscope
Currently our institute owns the following:
3 large Reichert "Z"-microscopes.
1 Leitz research microscope.
With the Reichert microscopes, a magnification of up to 3750x is easily attainable because of the slanted binocular [adapter] tube, which increases the normal magnification by 50%. If one uses a special Leitz objective 150 X Apochromat in conjunction with a 25x compensating ocular and the slanted binocular tube, a magnification of up to 4500x can be achieved, though not without great difficulty.
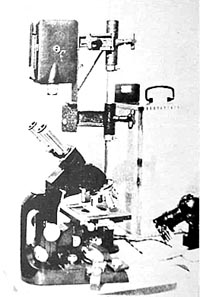

Figure 2. Setup for microelectrical studies.

Figure 3. Setup for long - and short-interval time-lapse photography.
Normally, examination is done under dark-field at a magnification of about 300x in order to study motility; for an overall appraisal of structure and type of movement, we usually use a magnification of 1200x; finally, at a magnification of about 3000x, the fine structure of the organisms and intracellular vibrations can be determined, which become visible only at this magnification. In order to assess accurately any movement inside the organisms, we also use a dark-field condenser, made by Reichert in Vienna, which enables us to do dark-field examinations at a magnification of about 3000x.

Figure 4. The two relay setups for the time-lapse photography.
This manipulation is extremely complicated and requires extensive preparation. A great many characteristic processes became observable only with the use of the Reichert "Z"-microscope; phenomena that in no way can be perceived with a vertical monocular tube or even with a nonslanting binocular tube. Therefore, verified results are hardly possible without the use of such optical instruments.
Microfilm Equipment
Every newly observed process that showed a typical characteristic was filmed immediately. Two kinds of film cameras were used: One was a CK Pan Film Camera, Kodak F 1.9, which permits a setting of 8 frames per second; i.e., a two-fold acceleration of movement. Normally, filming was done through a monocular tube at magnifications ranging from 300x to 1500x by positioning the film directly above the ocular of the microscope. We also succeeded in filming slow-moving structures at a magnification of 2300x through a slanted binocular tube; we did this by attaching the camera at an angle on to one of the oculars.

Figure 5. The microfilm apparatus and motor for the time-lapse photography.

Figure 6. Relay apparatus for long-interval time-lapse photography.
For time-lapse filming of developmental processes we used a large Cine Kodak Special Camera (F 1.9). This camera has the advantage of permitting individual shots and a very exact regulation of both light intensity and exposure time.
For time-lapse photography, two pieces of equipment were used: One was a time-lapse apparatus (Electric Release Control for Cine Kodak Special, produced by Eastman Kodak Co.).

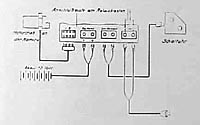
Figure 8. Circuit diagram for the long interval time-lapse apparatus.
By setting various relays, accelerations of movement are possible in the following order of magnitude:
4-fold acceleration (4 frames per second)
8-fold acceleration (2 frames per second)
16-fold acceleration (1 frames per second)
32-fold acceleration (1 frame per 2 seconds)
48-fold acceleration (1 frame per 3 seconds)
64-fold acceleration (1 frame per 4 seconds)
80-fold acceleration (1 frame per 5 seconds)
96-fold acceleration (1 frame per 6 seconds)
With about a 96-fold acceleration, [the frames contained on] one meter of film are exposed for 13 minutes 12 seconds. This camera is used for filming developmental processes and movements that are still visible to the naked eye, though with some effort and at high magnification.
To film developmental processes and movements that are no longer visible, we used a time-lapse apparatus furnished by Askania in Berlin.

Figure 9. Short interval time-lapse apparatus.
The switch and relay board were adjustable for the following acceleration speeds:
One frame in 15 seconds (240-fold time-compression)
One frame in 20 seconds (320-fold time-compression)
One frame in 30 seconds (480-fold time-compression)
One frame in 40 seconds (640-fold time-compression)
One frame in 1 minute (960-fold time-compression)
One frame in 5 minutes (4,800-fold time-compression)
One frame in 10 minutes (9,600-fold time-compression)
One frame in 15 minutes (14,400-fold time-compression)
One frame in 20 minutes (19,200-fold time-compression)
One frame in 30 minutes (28,800-fold time-compression)
One frame in 40 minutes (38,400-fold time-compression)
One frame in 1 hour (57,600-fold time-compression)
One frame in 2 hours (115,200-fold time-compression)
One frame in 5 hours (288,000-fold time-compression)
One frame in 10 hours (576,000-fold time-compression)
The last adjustment means that one meter of film could be taken over [a period of] 55 days and nights. The magnification in time-lapse filming ranged from approximately 300x to 1200x.
For the microelectrical studies, we used an apparatus especially built for this purpose (see Figure 2). A solid, round rod, with a movable cross-bar, is placed upright on a fixed base. This cross-bar, in turn, has two [vertical] glass tubes movable in two directions; [each glass tube has] a copper wire threaded through [the top end and connected to] a fine platinum wire projecting from [the bottom] end. The platinum wires are fastened to loops that are attached to two opposite sides of a trough on a stand. This [electrode] apparatus is connected to a Pentostat (manufactured by Siemens of Berlin) that makes exact measurements and permits a current adjustment as low as 0.2 mA.
Later, all filming was done with the aid of an optical attachment that permitted observation during the process of filming. The camera can be set up vertically above the ocular, as well as horizontally. By the Summer of 1937, we had produced the following: one complete film of preparation 6, bion experiment, and a film nearing completion: preparations 1, 2, and 3 (preliminary life forms, from swelling earth, coal, and soot). The laboratory is also equipped with all the necessary film-developing apparatus.
For electric potential measurements we used an ocillograph connected to a 3-tube D.C. amplifier. This instrument was produced by the Universiaetsinstrumentfabrik in Lund (Figures 10 and 11).

Figure 10. The 3-tube amplifier and silver electrodes (non-polarizable and unshielded).

Figure 11. Photographic film recorder (part of the set-up shown in Figure 10).
For the bacteriological studies, we set up a specially equipped laboratory complete with autoclaves (sterilization at 120 degrees) and dry sterilizer (sterilization up to 190 degrees).
Completed in October, 1937.
Signed: Wilhelm Reich
Part I
The Experiment
Chapter I
The Tension-Charge Formula
In this paper, I should like to present my observations vis a vis experiments I performed on the transformation of non-living matter into bacterial organisms. First, a brief discussion of the theoretical background:
During the course of approximately fifteen years of clinical work, I had come up with a formula for the function of the orgasm, which was later verified experimentally. [Footnote 2] In the autonomic life-support system, a sequence occurs whereby mechanical filling [turgor] or tension leads to an electrical charging [of the organism]; this is followed by an electrical discharge, which in turn results in mechanical relaxation.
This raises two questions:
First, does this formula apply only to the function of the orgasm or does it govern autonomic life functions?
Second, since the orgasm is a basic life phenomenon, this formula had to be manifest in even the most primitive biofunctions, such as the life activity of protozoa. I therefore postulated that the orgasm formula is identical to the life formula. Originally, I had no great hopes of proving the accuracy of this hypothesis within a short space of time. By accident, however, I solved the crux of this question quite quickly and consistently.
Furthermore, my clinical and laboratory experience had generated a wealth of ideas that served as guidelines for the biological research. Electrical experiments on the erogenous zones showed that vegetative excitations [of the organism] are functionally identical to their corresponding directions of electric flow. On the other hand, vegetative organ sensations appeared to be functionally identical with the onset of vegetative movement, which falls into two basic groups: The sensation of largeness and well-being, i.e., expansion, coincides with the onset of elongation as clearly seen in penile erection. Anxiety and sensations of unpleasure, on the other hand, are identical with the onset of a creeping back into the self, i.e., a contraction of the biological organism.
The change from expansion to contraction became obvious to me in my observation of ocean mollusks. The emission of electrical energy that occurs when this electric marine organism contracts confirmed my assumption that the sudden contraction is functionally identical to an electrical discharge. Therefore, I felt justified in leaping ahead to postulate that, on the one hand, electrical charging at the periphery, expansion, and well-being are functionally identical, as are electrical discharging at the periphery, contraction, and fright or anxiety on the other hand.
I postulated further that during expansion the distance between parts increased due to swelling. This must be closely related to an increase in electrical potential. Conversely, there is a decrease in distance between parts during contraction die to a loss of swelling, thus a greater tissue resistance, and a decrease in electric potential, i.e., discharge. Consequently, the physical [phenomenon of] electrical charge had to be felt directly as vegetative excitation.
Furthermore, I had come across the orgasm reflex about three years ago in my clinical work with neurotic patients who had hypertonic musculature. After removal of the muscular armor, separate vegetative spasms in various parts of the body combined in a unitary reflex involving the entire organism, which I called the orgasm reflex. It is the same phenomenon as the involuntary vegetative convulsions that occur at the acme of sexual gratification. From that I had to conclude that the autonomic nervous system expands and elongates during pleasure, but contracts in anxiety. The determining factor seemed to be the unitary functioning of the organism as a whole; that is, the ameba continues to exist in the metazoa in the form of a contractile and expansive vegetative life apparatus.
According to this view, the nerves within the organism no longer appeared to initiate the impulses, but merely functioned as organized conductors of vegetative impulses involving the body as a totality. The literature contained many references to the view that the ganglia in the autonomic nervous system function like storage batteries; and further, that the muscles represent the discharge apparatus, [with the discharge] resulting in movement. The body fluid, which in humans comprises about 80% of the total body weight, had to be regarded as the primary means for transmission of the electrical excitation.
The basic functions of the living, expansion and contraction, govern life; but they themselves consist of individual physical functions that are organized in a complex way. I will later discuss the evidence for this offered by colloidal chemistry. Here I should like to limit myself to a brief discussion of a pattern of uniformity, not only within the realm of the living, but also between organic and inorganic functions. As I mentioned, these were merely considerations that had developed from a great number of clinical and experimental investigations.
The biological directions "toward the world," represented by expansion, its opposite, "away from the world," "withdrawal into the self," represented by contraction, seemed to be reflected in the primitive model of a pig's bladder [as exemplified in] the mechanical process of expansion. When a pig's bladder is filled with air, it enlarges mechanically. The surface becomes tense and strives to restore the original condition, like a spring that is being stretched. This is opposed by an internal pressure exerted by the air. There are now three possibilities:
The internal pressure is lower than the surface tension; the bladder can be enlarged further without bursting
The internal pressure is equal to the surface tension; the bladder assumes a spherical shape and is stable.
Finally, the internal pressure of the air is greater than the surface tension; the bladder bursts
in the vegetative realm, an increase in internal pressure leads to contraction, as in the urinary bladder, or to constriction and division, as in the cell.
In electricity, charge and discharge are obviously separate processes. In inorganic processes, mechanical tension and relaxation, and electrical charge and discharge, are separate functions. In organic processes, on the other hand, these two physical functions are involved in a specific sequence" tension-charge-discharge-relaxation. This is the formula for vegetative functioning.
In chemical processes, there are substances which can produce tumescence, i.e., tension, and detumescence, i.e., relaxation. When KCI and lecithin act on tissues, the surface tension increases because of swelling; with calcium and cholesterin, the surface tension decreases due to detumescence and shrinking.
Now, it could not be without significance for an understanding of the function of the orgasm that tension and relaxation, tumescence and detumescence, expansion and contraction, charge and discharge, etc., are an integral part of the function of the parasymapthetic and sympathetic nervous systems, where they converge as one unified functional system. In a special investigation, "The Basic Antithesis of the Vegetative Life Functions" (1934), [Footnote 3] I had pointed out this relationship based on experimental results of other authors. Potassium acts like lecithin; lecithin like the parasymapathetic nervous system; finally, the parasympathetic nervous system entails pleasurable excitation, swelling, increased turgor, increased surface tension, and, as was finally shown, charge as well. On the other hand, calcium, cholesterin, the sympathetic nervous system, and unpleasure or anxiety, form a functional unity recognizable by the following processes: shrinking, contraction, discharge, and a decrease on surface tension.
I summarized these comparisons in a table four years ago as follows:
| Vegetative group (mutual comparison) |
General effect on tissues |
central/peripheral |
| Sympathetic
Calcium (group) |
Decreased surface tension. Dehydration. Striated muscle: flaccid. Decreased electrical excitability. Increased O2 consumption. Increased blood pressure.
|
systolic/vaso- constriction. cardiac stimulation. decreased peristalsis |
| Parasympathetic
Potassium (group)
|
Increased surface tension. Hydration (tissue turgor). Tetanic contraction of muscle. Increased electrical excitability. Decreased O2 consumption. Decreased blood pressure.
|
diastolic/vaso- dilation. cardiac relaxation. increased peristalsis.
|
Whatever had been described hitherto as "purposeful activation," "entelechy," etc., in biology, especially in its metaphysical aspects, seemed to be explained by just this leap from individual physical functions to their unified functioning; this is what directs the life process. Metaphysical principles of biology could thus be replaced by a dialectic materialistic interpretation of life processes.
The coordination of the life functions is controlled by the process of tension-charge within the individual organs, as well as within the entire organism. The concordance of inorganic and organic processes was evident in the function of tumescence-detumescence and charge-discharge.
The difference between the living and nonliving is a result of the specific combination of functions that otherwise occur separately in the nonliving.
Based on these postulates, summarized here, I now approached the biological experiments described below.
Chapter II
The Bions as Preliminary Life Forms
The vegetative streamings that I observed during my character-analytic work and the bioelectrical experiments induced me to investigate them further in protozoa under the microscope. The Botanical Institute of Oslo placed a protozoon sample at my disposal. I intended to prepare such samples myself and, therefore, inquired about the technique for culturing such preparations; I did not want to rely on information that I had acquired two decades ago in my biology laboratory courses and subsequently forgotten. Although I was familiar with the Leeuwenhoek infusions, I was surprised to learn that nothing else was needed except hay, i.e., half-dried grass, infused in water. I also learned that amebae are often found on leaves that have been lying in stagnant water for a long period of time. I was ashamed of the gaps in my knowledge of biology when I asked naively how these organisms got into such an infusion and was told that there were "life germs" everywhere, from which protozoa developed. It was obvious that I had deliberately, though unconsciously, "forgotten" the "germ theory." I specifically wanted to study ameba cultures in order to become familiar with the plasmatic streamings described by Hartmann and Rhumbler, which played such a crucial role in my theory of vegetative functions. However, there were only a few scattered amebae in the infusion I had received. It became apparent that it was not that easy to prepare new cultures made of amebae. In [response to] my distress, the assistant at the institute suggested that I prepare my own hay infusions and examine them "after approximately 10 to 14 days." I would then have no difficulty in finding amebae.
My knowledge of protozoology was limited. Nonetheless, I felt I could attempt to enter this field that was so completely new to me because of my thorough theoretical knowledge of biology and my background of clinical and research experience in orgasm function gathered over the past years. For the time being, I deliberately refrained from reviewing the biological literature so that I could be unbiased in my observations. I had an assistant compile the information found in the literature.
1. Vesicular Formation in Swelling Grass Blades
In the preparation, one could easily observe paramencia, various kinds of amebae, and, also, among other things, vermiform, undulating organisms. Vegetative plasmatic streaming was immediately apparent. The leisurely, undulating, slow-motion movements and plasmatic streamings were easily recognizable as those processes of tension: charge I was looking for. As a test, I passed a current of 0.5 mA through the preparation, and observed the same effect as had been described by Rhumbler and Hartmann, namely, that plasmatic streaming occurs at a faster rate when low currents are sent through. The amebae moved faster; granular movement inside the cell, as observed at a magnification of approximately 1500x, became extremely vigorous. The paramecia, on the other hand, seemed to become chaotic; their rapid, steady locomotion ceased; they began to rotate; it looked as if each current had a shocking, "traumatic" effect. When current was sent through for a longer period of time, approximately three minutes, the entire field became motionless except for the amebae. I increased the current to about 1.5 mA; now, the amebae took on a spherical shape, and even they ceased moving.
These observations seemed to be important, but I could not yet correlate hypothetically the current that was passed through and the resulting change in plasmatic streaming. However, there was another angle from which to examine the tension-charge formula.
It was obvious that living tissues absorbed fluid so that the membranes stretched mechanically in the process of swelling. Basically, the so-called tissue "turgor," excess accumulation of blood in the blood vessels, hyperemia, is really nothing other than an increased absorption of fluid (blood, lymph, water). I therefore concentrated on observing the changes that were taking place along the margins of the plant fibers.
During the course of about two days, the bright green fibers became blanched. In the solution there were many green chloroplasts. But there were strange changes taking place where the plant fibers were blanching and disintegrating. The previously striated and cellular tissue was transformed into vesicular structure. At the same time, the margins protruded in several places, mostly in the form of hemispheres, but also in jagged lines.
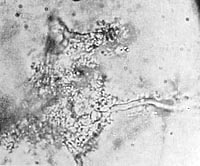
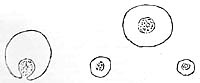
Obviously, the tissue fibers had previously absorbed water; they had undergone swelling; the chloroplasts had become detached; and a vesicular disintegration had taken place within the cellular structure of the tissue. In the previously clear solution there were heaps of vesicles everywhere, without membrane, motionless. They had the exact same appearance as the margins along the plant fibers that had undergone vesicular disintegration. The vesicular structure differed from one place to another. Here it was irregular, without a clearly defined membrane; there it was completely regular with a surrounding membrane that would luminate brightly when I turned the micrometer screw. Wherever there were distinct membranes, it was as if the unit had been "molded"; the more extensively the membrane had formed, the more taut the structure appeared. Gradually, the vesicles detached themselves from the fibers and floated around in solution. At that point, the plant fibers looked like branches that had shed their leaves (Figures 12, 13, 14).
I now followed the changes that were taking place not only in the vesicular structure but also in the membrane formation of one of these unite over a four-hour time period. At the margin of a piece of plant tissue there was an irregularly organized, membraneless, vesicular structure. It gradually swelled up, became larger, and detached itself from the plant tissue. Along the edged, fragmented double membranes appeared in increasingly extensive and distinct formation. The vesicular structure became more regular, more homogenous; the vesicles refracted the light with greater contrast. In its organization, the structure could hardly be distinguished any longer from a passing ameba. It assumed an oval shape and became more taut the more complete and distinct the membrane became. Inside, the vesicles were motionless, but the whole mass had assumed "shape." The structure disappeared when I tried to add more water to the preparation.
 F
F
Figure 12. Grass blades showing vesicular disintegration. Film prep. 8,700x.
Figure 13. Vesicles within a grass blade, 1500x.
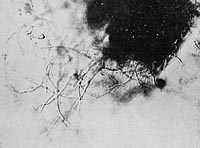
Figure 14. Plant fibers after detachment of vesicles.
Film prep. 8, 700x.
Detail of Figure 14
An ameba that was slowly moving along came close to the edge of the cover slip, dried up, and assumed a spherical shape. Now it could no longer be distinguished from the many heaps of vesicles that were scattered around.
Another observation pointed in the same direction: One can observe entities that exhibit the described vesicular structure, including a distinct membrane, that are attached to the fiber by a stalk. Occasionally, one can see such a structure struggling to tear itself away. It bounces away from the attachment, but is pulled back again by the threadlike stalk. Most importantly, it is distinguished from the lifeless structures only by its motility, while shape and content appear to be identical. Similarly, the many vesicularly-organized structures that move rapidly through the field, as if they were floating, appeared to be of the same type.
Let us summarize our observations:
1. Swelling plant fibers undergo vesicular disintegration.
2. Dried up amebae have a vesicular organization identical to the heaps of
vesicles.
3. Membrane-enclosed vesicular structures actively break away from the
disintegrated plant tissue.
4. The distinct membrane that is developing gives the heap a definite shape and tautness.
5. Many rapidly moving cells show a distinct membrane and a regular, vesicular structure.
Inevitably, this led to the surprising postulate that the vesicular ("honeycombed") living protoplasm of the ameba was very closely related to the vesicular heaps of vesicles resulting from the disintegrating plant tissue. Could it be possible that an ameba or other vesicularly structured Protista were nothing else but heaps of vesicles held together and shaped by a membrane?
Further observations and experiments speak best for themselves.
2. The Formation of Organisms Resulting from the Transformation of Grass and Moss Tissues into Life-like Structures
With the use of a certain species of grass for the infusion, the kind of Protista described is produced from disintegrating plant fibers through a series of preliminary stages. Immediately after the infusion is prepared, the grass blade shows a distinct margin with several claw-shaped spikes (Figure 15). During the course of approximately 24 to 28 hours, this margin undergoes vesicular disintegration as described (Figure 16). On a out the third day, I was able to see vesicular structures along the edge of the blade at irregular intervals, ranging in shape from round spheres to ovoid configurations, containing granules and vesicles of different sizes (Figures 17, 18, 19). The vesicles within the large membrane-enclosed vesicular structure cannot be distinguished from the vesicular configurations along the margin of the grass blade. The large, round structures are attached to the margin by a pedicle. Some of them do not change their form, but others display a strange behavior. The spherical heap which points away from the plant margin gradually elongates; this results in an opening at the anterior end, which is surrounded by very delicate threadlike fibers. The structure remains in this elongated position for about one to three seconds, until it contracts and suddenly resumes a spherical shape. These structures remind one of round apples or elliptical olives hanging from a branch. At the same time, or perhaps a little later, one sees them moving around in the solution, partly free and partly still connected by a pedicle to fragments of plant tissues that have become detached. The movement described is the same. One can observe the elongation followed by contraction to the spherical shape over many hours. Occasionally, one can see such a structure move away from the plant tissue as the coiled stalk is elongated. During contraction, many of them tend to bounce back against the plant tissue to which they are attached.
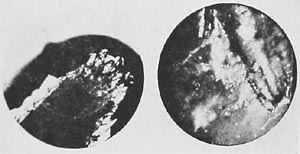
Figure 15. (left) Distinct grass margin. Microfilm prep. 8. Half dark field, 300 x.
Figure 16. Grass showing vesicular disintegration. Dark field, 1000x.
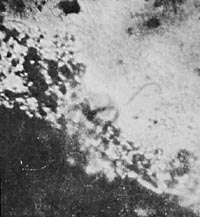
Figure 17. Vesicular formation along margin. Film, prep. 8, 1200x.
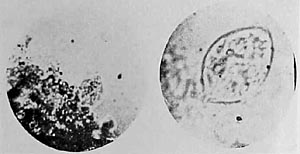
Figure 18. (left) Structured, membrane-enclosed configuration. Film prep. 8, 700x.
Figure 19. Oval, organized heap of vesicles, motionless. Film prep. 8, 1500x.
Now, can this already be called an "animal" or is it still a "piece of plant tissue"? how incorrectly the question is phrased. It is a transitional form, for, two days later, the same preparation showed the same organisms, without the plant margin, free in solution; however, at this time, they already had somewhat changed characteristics and could be seen in all different stages of development. There were still vesicular, spherical structures that were connected to plant fibers and that were expanding and contracting. Others carried out identical movements while already detached from the plant tissue. A third group was extremely unusual. The individual structures no longer resumed a spherical shape upon contracting, but retained their elongated form. In doing so, the 'mouth" was wide open, and they resembled paramecia. They moved like the latter, rapidly swimming around in the preparation in all directions; inside one could see a pulsating vacuole at the anterior end and vesicles in a circulatory movement. Those structures that repeatedly resumed a spherical shape showed the process of ingestion with special clarity during elongation. When the structure was completely elongated and the orifice was wide open, one could see the individual vesicles in the solution stream toward the orifice and then through it to the interior.
After two or three seconds, the orifice closed, the creature contracted, resuming a spherical shape, and now the vesicles began to move actively within; however, I could not yet pinpoint their movement in detail. Thus, it was not that the elongated structure moved toward the vesicles in solution and ingested them; rather the creature remained stationary and the individual vesicles streamed into its "mouth." It appears to be an electrical phenomenon. The creatures proved to be negatively charged, just like the vesicles. The accompanying photographs show various stages of development of these perhaps familiar creatures. We can see structures in spherical and elongated form; we can further see identical structures attached to plant fibers by pedicles. Some of these plant fibers clearly show a granular appearance and vesicular disintegration (Figures 17-25).
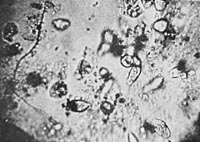
Figure 20. Org-protozoa on a blade of grass.
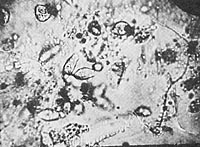
Figure 21. Org-protozoa on a blade of grass. (Figure 20 and 21 show two stages of development).
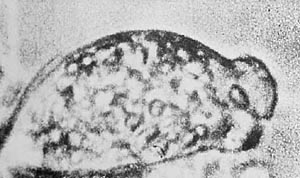
Figure 22. Org-protozoa on the grass margin. Vesicular structure of the plasma. 3000x
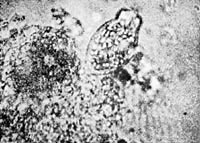
Figure 23. Completely structured heap of vesicles on grass blade. Film prep.8.
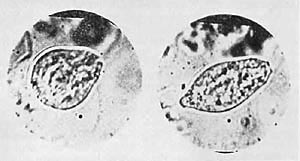
Figure 24. (left) Org-protozoon, egg-shaped, 1500x.
Figure 25. Same creature elongating.
The rhythmic alternation between spherical and elongated form seems especially important. Indeed, we may regard elongation as a result of the swelling process, during which the surface area becomes charged. It denotes the direction "away from the self, toward the world." The intake of solution and vesicles through the open "mouth" possibly signals a process leading to assumption of a spherical shape following contraction. We may regard contraction as a process of discharge, just as muscle spasms follow from electrical stimulation. When weak currents are sent through the preparation, contraction occurs prematurely, before the structure has completely elongated. The detailed relationships are still to be clarified. To progress in this work, it was necessary to postulate that here the tension-charge-discharge-relaxation process is directly visible. Of course, there are vital questions that remain unanswered, such as what enables this creature to retain its elongated shape and how does it lose this property to return to a spherical form? These observations have produced evidence that at least this type of protozoon develops from the transformation of plant tissue through a series of developmental stages. It would be meaningless to categorize or classify them at this time. The observed preparation was recorded on film. (Prep. 8). The contractile structures were named "Org-protozoa" (after "orgastic" convulsion).
3. Evidence for the Vesicular Nature of the Flowing Amebae
The first hypothesis, that the flowing amebae were vesicles enclosed in a membrane, was formulated about two years ago. However, the study of earth bions largely deflected us from the questions surrounding the amebae. Evidence for this hypothesis was not forthcoming. Controlled microscopic observations were lacking. I now hoped for a time-lapse film apparatus that would disclose the process on film. However, for a whole year, the problem could not be solved because of technical difficulties. Time-lapse photography requires a totally motionless preparation and an unstirred solution. It soon became obvious that grass infusions die from lack of oxygen when enclosed in a paraffin. However, a film of the membrane formation around the vesicle and the subsequent development of amebae from those structures could constitute the major evidence.
The technical problem was solved in the following manner: Two to three pieces of moss or grass, clearly separated from each other, are placed on a hanging drop slide, and water for the preparation is added. A piece of was is put under each of the four corners of a cover slip, which is then placed over [the slide] in such a way that no air bubbles form. The cover slip is now attached firmly with paraffin to the two longer sides of the slide. In doing so, one has to make sure that approximately one fourth of the slide cavity is not covered by the cover slip. Water is then pooled along the two sides of the cover slip not covered with paraffin so that the evaporating solution can be replaced continuously with water from the two reservoirs set up o either side. About every two hours, the two reservoirs have to be replenished with water. In this way, the technical problem is solved, and time-lapse photography is made possible over several days; however, some effort and attention are necessary so that there is no delay in refilling the reservoirs lest the preparation dry up in the meantime. With the aid of these technical considerations, dried moss in water was shown to produce amebae, in addition to other protozoa, under conditions not yet fully understood. Thin fragments of grass, which have been made to swell, disintegrate into the most delicate vesicles. One can then observe them growing larger and [note] a distinct membrane being formed around aggregates of vesicles which are initially spherical. When the membrane has reached a certain size, here or there it begins to protrude, but the whole structure us still motionless. A small, round circle appears at the center. Then, a loose outer layer is formed around the large membrane-enclosed structures that have at their center a small, almost punctate vesicle. This will later become the ectoplasm, while the vesicle within the large, round structure give rise to the coarse, vesicular quality of endoplasm.
After several hours, the structure becomes detached from the fiber and continues to move like an ameba. Only in the very last stage of this process does streaming occur in the protruding membrane. The detachment takes place gradually. When the structure has broken free from the fiber, one can already see pseudopodia in various parts of the organism. In this way there is a continual development of amebae. Sometimes, there are two or three simultaneously forming within a short period of time; at other times, they form at specific intervals. The detached amebae divide. Often, they are found in colonies. In this way, they can hardly be distinguished from the margins along moss fibers which have undergone swelling. Evidence for the formation of amebae from swelling moss fibers was provides by this direct observation. (Compare the microphotographs from the time-lapse film, Figures 26-31.)

Figure 26. Moss beginning to swell, 300x

Figure 27. Moss showing vesicular disintegration. Dark Field, about 800x.
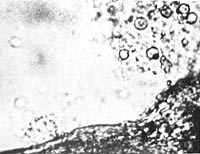
Figure 28. A phase in the development of flowing amebae. The vesicles directly above are from swollen moss.
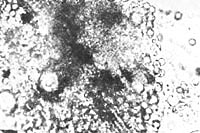
Figure 29. A further stage of the development of the ameba in the same preparation. The vesicles on the left are in the process of dispersing. 1000x
Figure 30. Fully developed amebae wandering off. (Directly over the dispersing vesicles.) 800x.
Figure 31. Vesicular structure of an ameba. 1500x
4. Motile, Vesicular Earth Crystals and Earth Bions
My observations and consequent postulates were at variance with the "germ theory." I purposely avoided orienting myself anew on the prevalent views of the "origin of life from life germs." After these first observations, more than ever, a totally unbiased approach to the work was necessary. For comparative purposes, I set up several unsterile infusions consisting of a tulip leaf, a rose petal, grass, and plain earth. After three days, the tulip leaf and rose petal infusions did not show any protozoa; the grass infusion was full of motile rods, vesicles, and various Protista; the earth infusion (Prep. 32) was a big surprise.
Immediately after the preparation of the infusion, microscopic observation showed distinct membrane-enclosed, unorganized crystals that were completely motionless (Figure 32).
Figure 32. Unorganized earth crystal. Film prep. 1,2,3 - 800x.
Scattered among the crystals, one could find vibrating vesicular structures. Here and there bright green, elongated rods without nuclei were slowly moving around. On the third day, the same open preparation looked quite different. It was full of motile structures which, though sharply angular, moved in exactly the same fashion as the rods and vesicles. It was noticeable that the small vesicular structures often adhered to the surfaces of rods or larger structures. The structures had a taut, vesicular organization. Many of them had changed in their organization; striations had appeared inside, which here and there had undergone vesicular disintegration. The vesicular configurations within the organized crystals could not be distinguished from the vibrating, freely moving vesicular structures in the solution.
On the seventh day, the vesicular disintegration and organization had progressed extensively. At a magnification of only 700x, one could observe clearly delineated protrusions along the edged of the angular, irregular, brow-colored clumps of earth, which looked like tubular vesicles alternately expanding and contracting; angulation was also observed. At a magnification of 1625x, I observed a brown clump whose margins had protruded here and there like vesicles. It was connected to another clump by a vesicular, striated, organized substance. At the junction the clump was bending and elongating. At first, I thought I had been mistaken, but continues careful observation left no doubt: The clump of earth moved as if it had joints, expanding and contracting.
Figure 33. Crystal showing the beginning of vesicular organization. Six week old prep. 1a, 1000x.
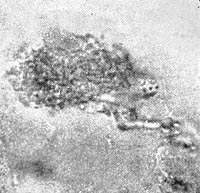
Figure 34. Crawling crystal showing vesicular organization.
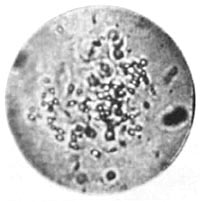
Figure 35. Moving pseudo-ameba. Sterile earth bound together by gelatin.
After another seven days, the process of vesicular disintegration and striated organization had progressed significantly, as had the vesicular disintegration of the crystal edges. The vesicular protrusions along the crystal edges showed three kinds of movement:
Rotation about the axis;
Expansion and contraction;
Angulation
Let us call the newly described structures on the crystals "plasmoids" (Figures 36, 37).
I passed a 1 mA current through, which I very gradually increased to 2 mA. The movements increased significantly: The vibrating motion of the vesicles became more rapid; expansion and contraction more distinct. When the current was turned on, the vesicles moved toward the cathode. Therefore, they were positively charged particles. When the current was turned off, this movement stopped as promptly as it had started when the current was applied. When the current was reversed, the direction of movement of the individual structures was quickly reversed. When current was sent through for a prolonged period of time, the swollen tubular structures along the edges attempted to break loose in a jerky fashion. Occasionally, these attempts continued for a short while after the current had been turned off. Often, the tubular structures doubled in length. Repeated checking of their reaction to electricity always gave the same result. Therefore, the electric current had some influence on the spontaneous movement, especially elongation, that could not yet be clearly defined.
I will return later to the above reactions, and focus now on some other phenomena observed during the subsequent course of my work.
Let us note that, so far, we have observed two phenomena, which are-in all likelihood-fundamentally different:
1. the vesicular disintegration of swelling plant fibers, i.e., organic tissues, and
2. the vesicular organization of earth crystals, i.e., inorganic material, with subsequent formation of motile, tubular structures and other motile particles.
From then on, I attempted to reproduce the two phenomena experimentally over an over again and to ramify the experimental setup. Above all, I wanted to verify my hypothesis that the vesicles formed from the swelling of substances were actually identical to the vesicular structures within the amebae.
I henceforth replaced the water in the preparation with 0.1 N KCl to test the effect of potassium on swelling. I found that earth treated with KCl began to swell more rapidly and the described phenomena appeared more distinctly; a similar result was obtained when completely- or half-dried grass was treated with KCl. In the grass infusions, I could regularly elicit the production of motionless, vesicular heaps. I became more and more accustomed to seeing the individual vesicles streaming in and out. It was also possible to reproduce over and over again the progressive vesicular disintegration of the earth preparations treated with KCl.
I was interested in artificially coalescing the vesicles formed as a result of the disintegration. I therefore added very dilute red gelatin to the grass and earth preparations that were in a stated of advance disintegration. After only a few hours, one could observe ameboid structures that had not existed previously and that were not present in the preparations lacking gelatin. It took about one to two days for these structures, which I dubbed "pseudo-amebae," to develop fully. They were heaps of vesicles which moved around jerkily in different directions. Occasionally they had individual rods protruding form their membranes like motile spikes. The movements were jerky, not organically flowing like cytoplasmic streamings of real undulating amebae. Aside from locomotion, movements inside the structures, as well as contraction and expansion, could be clearly seen at a magnification of approximately 2000x. As I had postulated, the gelatin had consolidated a number of individual vesicles into heaps of vesicles. The heap of vesicles now continued to move as a unit, the next problem was to determine how this movement originated. Flowing cytoplasmic streaming, observable in many amebae, could not be seen in these preparations (Figure 35).
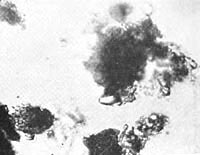
Figure 36. Plasmoid earth crystal, motile protrusions along edges, 1650x.
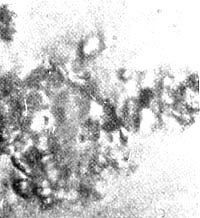
Figure 37. Motile plasmoid earth crystal.
Figure 34 shows a crystal completely organized into vesicles and connected by a stalk to a second crystal (organized only along it left margin). At the time the photograph was taken, this crystal moved very slowly in the direction of the arrow, pulling the partially structured crystal behind it.
Now, one could argue that the pseudo-amebae described above had not been assembled artificially, but that "germs of living organisms" had invaded the unheated, open preparation. In order to test the validity of this objection, I now began to heat the preparation in sealed glassware for a quarter to half an hour.
This trial brought a totally unexpected result: It became apparent that the heated infusions displayed far more abundant an active life immediately after their preparation that did the unheated ones after days of swelling.
In the unheated preparations, the vesicles were scattered individually; unorganized crystals were predominant; the vesicular organization of the crystals developed slowly over weeks. The heated earth preparations, on the other hand, showed-immediately after heating-a vast number of tightly packed, active, continually moving, spherical as well as irregular-shaped, vesicles. In the unheated preparation, the solution usually remained clear and contained sediment. In the heated preparation, it immediately clouded up and became colloidal. Electrical testing of the heated preparation showed that the vesicles were also positively charged particles that moved toward the cathode when a current of 0.5 and 1 mA was passed through. On subsequent addition of diluted sterile gelatin, the same result was obtained as earlier with the unheated earth: the aggregation of Individual granule-vesicles into motile ameboid structures. The same result obtained when we heated a mixture of earth, KCl, and gelatin.
However, I was greatly puzzled by the presence of motile structures in a heated, tightly closed preparation immediately after heating. How was it possible that the heated preparation showed much more life than the unheated, unsterile, open one? This seemed to contradict all laws of sterilization. Six days after heating, a tightly closed preparation showed, at a magnification of approximately 1500x, that most of the structures exhibited the familiar movements. There were also plasmatic protrusions and formations, brightly luminating spots with three or four threadlike or rodlike extensions.
Most of the crystals were completely composed of vesicles. Corresponding controls with fresh, unheated earth and water preparations showed the same result over and over again, namely, a paucity of vesicular structure and a dearth of vesicles lacking a nucleus. In my observations, I had to learn to distinguish the spontaneous movement of clumps of earth that had undergone swelling from the passive movement of those recoiling from collisions with motile structures. I could no longer have any doubts about the far more lively nature of the heated preparations; nor the incomparably more abundant formation of contractile sacs and swelling along the edges.
With the use of strong lenses and water immersion and a magnification of approximately 2300x to 3000x, I could clearly observe pulsation in separate parts of the contractile sacs. Solidly crystallized structures with organized margins, which seemed to be connected to one another to one another by a gelatinous substance, attempted to "break free." I was surprised to see, though only at the highest magnification, green, luminous highly motile formations within the gelatinous substance, adhering to some of the structures and indistinguishable from the green, luminous, nucleus-containing rods floating freely in solution. This led to the assumption that the clumps of earth underwent swelling and developed into vesicles or rods in certain places. There are as yet unknown factors involved that determine whether these swollen units remain inside the crystal or break looses and float freely in the solution. When an electric current of 0.5 to 1 mA was passed through, these preparations also showed the now familiar phenomena of vesicular protrusions, increased vibration of the vesicle against each other, movement toward the cathode, etc.
It became increasingly clear to me: The more vesicular the structure of the crystal, the more motile it becomes. In order to solidify my future work, I had to postulate that the vesicular components are swollen units of matter of high electric charge, which are held together by a gel-like substance, and which move within the ameboid structures just as they do in their free state as separate vesicles without a nucleus.
Repeated control experiments with heated earth gave the same results over and over again: motile vesicles, structures with and without a nucleus, pseudo-amebae, and divisions.
In order to rule out faulty preparations as the source of these phenomena, I set up control infusions of unheated earth, as well as tree and tulip leaves, etc., which I left exposed to the air. Over and over again, the above described result was obtained, namely, that the unsterile preparations shoed far less vesicles than the heated ones, and that it was far more difficult to obtain marginal layers that had become motile or had undergone vesicular disintegration.
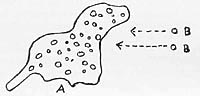
The "process of ingestion" drawn from life.
Ameba A did not move toward bions B; however, after a certain distance, the bions moved rapidly toward the ameba and disappeared inside (note direction of arrow)
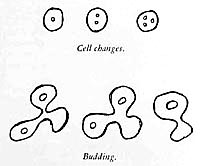
The question now arose whether these structures, if they actually represented living matter, showed, aside from movement, contraction and expansion, still other known properties [of the living], such as cell division. Therefore, I began to study the earth-KCI-gelatin preparations very carefully. Within a short space of time, after focussing for hours on a single field or structure, I could observe pseudo-amebae divide.
The more familiar I became with the preparations, the more probable it became that I was dealing with living organisms which, however, lacked completeness, so to speak; for example, the movements of the pseudo-amebae were jerky, sluggish, vibrating, without inner streaming, "mechanical." The structures had to be preliminary forms of living organisms. To begin with, for my own personal use, I called them bions. Could full life develop from these bions?
It was very satisfying to detect the function of "ingestion," in addition to cell division. I was able to observe that such pseudoameboid heaps of vesicles ingested individual vesicles that were freely moving about. I thus had to free myself of an idea that is erroneously connected with the word "ingesting." No doubt, we make the mistake of using anthropomorphic expressions when describing protozoal structures. When we say "ingesting," we automatically think that a "rational" living organism "takes in" something from its environment "in order to maintain its life." Only after I had freed myself from this erroneous thinking was I able to incorporate the phenomenon of "ingesting" logically into my other observations: To begin with, it seemed inevitable that the cells I had in front of me were composed of vesicles and rods, just as a multicellular body is composed of individual cells. This contradicted the view that cells are the basic units of living matter; because then it would not be the cells but the vesicles that would constitute a biological unit, and the cells would already be complex structures. Yet, these vesicles and rods, as shown by their reaction to electricity, were highly charged, tautly stretched structures. Hence the obvious assumption that a heap of vesicles carries out its "process of ingesting" by attracting individual vesicles which are free-floating and electrically charged, and that it incorporates them into the aggregate it is forming, just as we had observed with the contractile protozoa.
This was simply a hypothesis that came to me. I would have gladly accepted any other workable one. At this stage in my work, further development of the experiments and their controls seemed to me most important. Over and over again I came up against the question as to how it was possible that heated substances showed more life than unheated, unsterile ones. Originally I thought that the rapid, vibratory movement of the vesicles after heating was due to heat. However, several tests with electric current showed this movement to be of an electrical nature, since this vibratory motion also increased when current was sent through. I did not yet want to consider whether I was dealing here with actual bacteria or cocci, or whether these were structures of a different nature that had been formed a s result of heating. Since it had become possible to produce pseudoamebae from heated earth, gelatin, and KCI, it seemed feasible to perform a further control experiment with other substances. According to my hypothesis, there had to be substances that removed the "incomplete" character from the structures.
Later, I proceeded to autoclave the earth in KCI at 120 degrees. The result was much better as compared to the merely heated coal (at 180 degrees), i.e., there were a greater number of more organized crystals, and the vesicles and heaps of vesicles were almost completely motile. I will return to this when I describe my attempts to culture them.
The observations described so far confirm the biological correctness of the tension-charge formula. The vesicular swelling represented mechanical tension; the occurrence of electrical positivity or negativity on the individual vesicles represented electrical charge. However, most of this still remained in the dark.
5. Albumen Preparations (Prep. 6)
Based on the tension-charge formula, the substances necessary to complete the [development of] function in the structures had to possess the property of swelling. Since, according to other studies, lecithin behaves like potassium and cholesterin like calcium (the former causes tumescence, the latter detumescence), I proceeded to add lecithin and cholesterin to the earth preparations. The result was as follows: In addition to the already described phenomena, there were a few others which had not been detected previously. First of all, it was noticeable that the preparation was full of tubular, regularly formed structures, which slowly changed their shape (Figures 38, 39, 40).
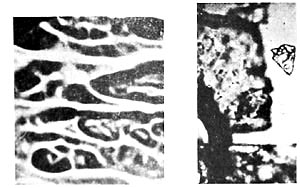
Figure 38. (left) Lecithin, spread dry on cover slip, 400x.
Figure 39. Cholesterin crystals, 400x.
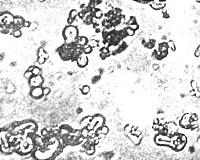
Figure 40. Tabular structures from lecithin. Lecithin in 0.1 N potassium chloride, 1200x.
Control tests involving identical treatment of each substance individually soon revealed that these structures had to be attributed to lecithin. They had the following characteristics: The tubular, often regularly notched structures elongated and thickened, bent, changed their position, and in particular showed budding; i.e., in some places, another tubular structure grew out of the original one, which again branched, etc. When I added only KCI to lecithin, I obtained the identical structures. They did not display any organic movement. The "growth" and budding observed I believed to be attributable to the intake of fluid, as into a sac. It ceased after approximately forty-eight hours. However, lecithin, without doubt, was capable of forming structures. The movement connected with this growing and budding was fundamentally different from that of the pseudoamebae.
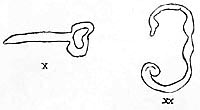
x Elongating tubular structure.
xx Luminating green expanding and contracting; when current is applied, elongation is towards the anode.
I now added chicken albumen to the earth-lecithin preparation. The result was beyond all expectations. In unheated preparations, round cells formed after a few minutes; they contained dark nuclei which divided frequently and in rapid succession. The cells divided either through a central cleavage furrow, through which the nucleus also divided, or by budding they produced smaller cells, which then detached from the mother cell. In some areas, one could see distinct movements of breaking away. What I am attempting to describe here briefly was unfolded in reality only after long, laborious observations.
Cell-like structures obtained upon addition of chicken albumen.
Nucleus-like formation inside tubular structure, and cell-like structures.
To the extent that I learned to observe and follow the phenomena, they became more consistent, for I gained confidence in identifying them. Thus, actual cells were formed upon the addition of albumen. Cell formation did not occur without the addition of lecithin and cholesterin; otherwise, only heaps of flaky albumen were observed in the preparation, as can also be seen when unboiled albumen is examined under the microscope. Neither did lecithin with water alone give rise to motile cells.
We also observed that in the unheated albumen preparations the formation of motile substances occurred much more slowly and sparsely than in the heated preparations. The latter could also be distinguished outwardly from the unheated ones by the colloidal turbidity of the solution.
The unheated preparations were far less cloudy and showed a greater amount of sediment. The colloidal particles formed after heating were therefore in suspension, which, according to all previous knowledge, must be attributable to their electrical charge. I let the preparations stand for weeks and was unable to observe that there was a tremendous increase in the number of individual formations. However, after a certain time, about six to eight weeks, the solution started to clear again and the particles sank to the bottom where they formed a thick, whitish layer. Microscopic examination showed that, when the colloids were no longer in suspension, they also lost their motility and their reaction to electricity. The structures were "dead." Let me recapitulate briefly:
Lecithin with KCI alone does not produce cells, but only various kinds of regularly formed tubular structures. Moreover, there is no organic motility, but only growing and budding which is obviously due to fluid intake. Adding only KCI to albumen does not result in cell formation. But albumen plus lecithin plus KCI plus cholesterin results in cell formation.
Figure 41. Bions immediately after their formation
+ = structures formed upon addition of coal bion prep. 6b, 1000x.
Figure 42. Fresh sterile bions, bion prep. 6c.
I now complicated the experiment by adding gelatin also. Therefore, I was now mixing together albumen, earth, lecithin, cholesterin, gelatin, and KCI. Addition of gelatin caused plasmoid vesicular structures to be visible immediately after heating; they moved vigorously and displayed a much finer structure and far more life-like vegetative motility than the pseudoamebae in the earth preparation described earlier. It was no longer jerky, no longer mechanical, but organically flowing. (Figure 41). The longer the preparation was heated, the more complex were the ameboid creatures obtained in this way; they possessed many more [life] functions than the heated earth preparations. While the pseudoamebae in the earth preparations simply moved from place to place, displayed minimal or no movement of the particles within, and showed only very weak and occasional streaming, contraction, and expansion; the ameboids in the albumen preparations showed streaming, internal movement, contraction, expansion, locomotion. The function of division was present in both, as was the function of "ingestion" (Figures 42, 43).
The ameboid structures in the albumen preparations showed a much finer vesicular structure. Also, after a few days, one could see structures with a homogenous plasma that showed flowing, pseudopodal movements. Also present were contraction, expansion, division, budding, "ingesting," locomotion. Still unanswered were questions of metabolism, culturing, staining, and the bacterial nature of the structures we had produced.
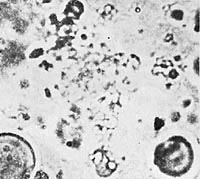
Figure 42. Vibrating amebae moving in place. Bion prep. 6. 2300x.
Basic evidence for the possibility of creating the living from the non-living is the fact that when various substances are used, the addition or omission of one or another substance can change the biological nature, the composition, and the activity of the motile structures.
It is not necessary to mix the substances in exact ratios. It is as if the structures form according to still unknown laws, as if the ration of substances that combine to produce motile life, motile forms, is governed by a self-activating principle. Here everything is in the dark.
Subsequently, I also added meat bullion, milk, and egg yolk as nutrients to the bion mixture. Also, carbon in various forms: finely pulverized coal sterilized dry; coal dust autoclaved in KCI; incandescent coal dust, KCI, dry or in bullion; finally, dry, sterilized or incandescent soot. The best results were achieved upon addition of soot; extremely small, but exceedingly motile amebae of the finest structure were produced. Bion mixture 6 is now prepared in various ways:
6a: Unsterile mixture;
6ab and 6ac: The unsterile mixture is heated (100 degrees, 1/2 hour)
or autoclaved (120 degrees, 1/2 to 1 hour);
6c: The substances are previously sterilized;
6cb: The sterilely produced mixture is heated afterwards;
6cc: The sterile mixture is autoclaved again;
6ccc: The sterile mixture is autoclaved twice.
The best culture results are obtained with mixture 6cc. Inoculation is done for the first time after three to four days.
Can one speak here about artificially created life? Vanity would like us to answer in the affirmative. But correct consideration leads to a no! If the living were a distinct metaphysical realm totally separate form the nonliving, then I would have the right to say that I create "artificial life." However, my experiments demonstrated that there are developmental stages ranging from the nonliving, nonmotile, to the living; that, in nature, the creation of life from inorganic matter probably occurs every hour, every minute. In this case, one can hardly talk about artificial life. I merely succeeded in uncovering the developmental process of life experimentally. Perhaps a new kind of organism was thereby also produced artificially.
On 1-8-1937, I sent the following report to Professor du Teil:
In lieu of a detailed report at this time, I should like to inform you of a thermal experiment based on the tension-charge formula. I am reporting here only the protocol and result of the experiment. Currently, a file is being made at the Institute for Sex-Economic Research which will facilitate an understanding of the discoveries. Likewise, samples of the colloid preparation will be distributed.
First, at the present experimental stage, I mix 100 cc of sterile Ringers solution with 0.1 N KCI solution. To this mixture I add dissolved red gelatin until a faint pinkish -red color appears. One tweezer-tip full of coal dust is added to this mixture, and the same amount of cholesterin crystals. The total experiment is based on the principle of adding together in a specific sequence those that are necessary for cell formation.
We then dissolve one teaspoon of fresh, clear chicken albumen in about 50 cc of sterile KCI solution, and add both to the previous mixture. After brief stirring, the albumen is dissolved. Microscopically, no structures, movement, or plasmatic forms of any kind are visible. At high magnification (1000x), we see merely the typically motionless coal and cholesterin crystals.
At this point, we add about 1 to 2 cc of milk and some egg yolk. The latter causes the previously clear mixture to become cloudy.
We now prepare a second solution: lecithin ointment is stirred into KCI solution. Under the microscope, at a magnification of about 5-900x, we can observe unusual structures forming and growing: tubular structures which are swelling, budding, bending. Inside the tubular structures, we see no organization at all. However, heaps of vesicle are found occasionally. Organic movement is lacking. They are the purely physical phenomena of swelling corresponding to changes between internal pressure and surface tension.
If lecithin solution is now added to the first mixture, we immediately see a progressively increasing cloudiness that is yellowish-grey in color. Under the microscope, the surprising picture of motile life unfolds: vibratory movement from place to place, budding, division of cell-like structures, jerky to flowing motion. The organic motility can be observed from 1500x, but is best seen at 2300x to 3000x with a binocular microscope. At a magnification of about 3000x, while focussing continually on one structure, one can see luminous vesicular, nuclear structures which are [constantly] changing form. Basically, one can distinguish four groups of active, motile structures: spherical, nuclear-like vesicles; rods; round, cell-like structures containing nuclei, which move in solution but do not show any movement within; and finally, ameboid structures. The latter are especially striking because they show movements of contraction and expansion within the organisms at a magnification of approximately 3000 to 3500x. Indeed, their locomotion is already visible at a magnification of only 2000x.
At least according to past and current experiments, the structures produced a negative electric charge; they move toward the anode. When the colloid mixture is allowed to stand for 6 to 8 weeks, a thick sediment is formed on the bottom; the solution becomes increasingly clear. Upon examining the sediment at the previously mentioned magnifications, it is shown that the described motility has been lost; the structures are "dead." I will report on the control experiments performed in my detailed discussion. Currently, I am working on tests concerning metabolism and sensitivity to stains. It is certain that the structures can be cultured; for when the mixture is heated until all the liquid has been evaporated and the solid matter is extracted, inoculation of this extracted residue with sterile, colloid mixture produces growth [of structures] which have already propagated to the fifth generation.
In animal tests, these structures, if kept sterile, proved to be harmless when injected subcutaneously into mice and guinea pigs.
The question as to whether we are dealing with complete, living organisms can only be answered in conjunction with other experiments that I will discuss at another time, and only after all tests and controls have been performed. The experiment described was recorded on film.
We called the artificial, lifelike structures "bions."
(To be continued)

{kind=link}